Regulacija sinteze holesterola
Regulacija sinteze holesterola - njegovog ključnog enzima (HMG-CoA reduktaza), provodi se na različite načine.
Fosforilacija / defosforilacija HMG reduktaze. S porastom omjera inzulin / glukagon, ovaj enzim fosforilira i prelazi u aktivno stanje. Delovanje inzulina se izvodi preko 2 enzima.
HMG-CoA reduktaza kinaza fosfataza, koja kinazu pretvara u neaktivno defosforilirano stanje:
Fosfotaza HMG-CoA reduktaza pretvarajući je u defosforilirano aktivno stanje. Rezultat tih reakcija je stvaranje defosforiliranog aktivnog oblika HMG-CoA reduktaze.
Zbog toga se tokom perioda apsorpcije holesterol povećava. Tokom tog razdoblja, povećava se i dostupnost početnog supstrata za sintezu holesterola - acetil - CoA (kao rezultat konzumiranja hrane koja sadrži ugljikohidrate i masti. Kako se tijekom razgradnje glukoze i masnih kiselina formira acetil CoA).
U postabsorbentnom stanju glukagon putem proteingenaze A stimulira fosforilaciju HMG - CoA - reduktaze, pretvarajući je u neaktivno stanje. Ovo je djelovanje pojačano činjenicom da istodobno glukagon potiče fosforilaciju i inaktivaciju fosfotaze HMG-CoA reduktaze, zadržavajući na taj način HMG-CoA reduktazu u fosforiliranom neaktivnom stanju. Kao rezultat toga, endogena sinteza inhibira sintezu holesterola u periodu postabsorpcije i tokom posta. Ako je udio kolesterola u hrani doveden na 2%, tada se sinteza endogenog kolesterola naglo smanjila. Ali potpuni prestanak sinteze holesterola ne dolazi.
Stepen inhibicije biosinteze holesterola pod uticajem holesterola koji dolazi iz hrane varira od osobe do osobe. Ovo ukazuje na individualnost procesa stvaranja holesterola. Smanjenjem intenziteta sinteze holesterola moguće je smanjiti njegovu koncentraciju u krvi.
Ako se poremeti ravnoteža između unosa holesterola s hranom i njegove sinteze u tijelu s jedne strane, i izlučivanja žučnih kiselina i holesterola s druge strane, mijenja se koncentracija kolesterola u tkivima i krvi. Najozbiljnije posljedice povezane su s povećanjem koncentracije kolesterola u krvi (hiperholesterolemija), dok vjerovatnoća za razvoj ateroskleroze i holelitiaze raste.
Porodična hiperholesterolemija (HCS) - ovaj je oblik mnogo češći - oko 1 pacijenta na 200 ljudi. Nasljedna manjkavost HCS-a je kršenje apsorpcije LDL-a ćelijama, a samim tim i smanjenje stope katabolizma LDL-a. Kao rezultat toga, koncentracija LDL u krvi raste, kao i holesterol jer ga ima puno u LDL. Stoga je za HCS karakteristično taloženje holesterola u tkivima, posebno u koži (ksantomima), u zidovima arterija.
Inhibicija sinteze HMG-CoA reduktaze
Konačni produkt metaboličkog puta holesterola. Smanjuje brzinu transkripcije gena HMG-CoA reduktaze, čime inhibira vlastitu sintezu. Jetra aktivno sintetizira žučne kiseline iz kolesterola, te stoga žučne kiseline inhibiraju aktivnost gena HMG-CoA reduktaze. Budući da HMG-CoA reduktaza postoji nakon sinteze oko 3, inhibicija sinteze enzima ovog holesterola je efikasna regulacija.
Niste pronašli ono što tražite? Koristite pretragu:
Izmjena holesterola
Fond holesterola sadrži slobodne holesterole i estere holesterola, koji se nalaze i u ćelijama i u lipoproteinima u krvi.
II deo Metabolizam i energija
U ćelijama se esterifikacija holesterola događa djelovanjem acil-CoA-holesterola-aciltransferaze (AChAT):
Acyl-CoA + holesterol - * HS-KoA + Acylchoterol
U ljudskim ćelijama uglavnom se formira linoleilhoterol. Za razliku od slobodnog kolesterola, njegovi esteri u staničnim membranama sadrže se u veoma malim količinama i uglavnom se nalaze u citosolu kao dio kapi lipida. Stvaranje estera može se s jedne strane smatrati mehanizmom za uklanjanje viška holesterola iz membrana, a s druge strane, kao mehanizmom za skladištenje holesterola u ćeliji. Mobilizacija resursa događa se uz sudjelovanje enzima esteraze koji hidroliziraju estere holesterola:
Acil-holesterol + H 2O - * Masna kiselina + holesterol
Sinteza i hidroliza estera događa se u mnogim ćelijama, ali posebno je aktivna u stanicama kore nadbubrežne žlijezde: u tim ćelijama do 80% svih kolesterola predstavlja estere, dok je u ostalim stanicama obično manje od 20%.
Stvaranje estera u liponroteinima u krvi odvija se uz sudjelovanje lecitin-holesterola aciltransferaza (LHAT), koji katalizira prijenos acilnog ostatka iz i-položaja lecitina u kolesterol (sl. 10.31). Brzina esterifikacije za različite lipoproteine značajno je različita i ovisi o prisutnosti apolipoproteina koji aktiviraju LHAT (uglavnom apo-AT, kao i CI) ili inhibiraju (C-II) ovaj enzim. Najaktivniji LHAT u HDL-u, u kojem je apo-AT morati Olee 2/3 svih proteina. Najveći coli počasti formirana estere oleinske i linolne kiseline. Druga lipoproteina estera formiranje javlja sporije nego u HDL.
Sl. 10.31. Stvaranje estera holesterola pod delovanjem LHAT-a
LHAT je lokaliziran u površinskom sloju HDL-a i kao supstrat koristi holesterol u fosfolipidnom sloju. Ovdje se formiraju esteri holesterola, zbog njihove potpune hidrofobnosti, slabo zadržavaju
Poglavlje 10. Metabolizam i funkcija lipida
fosfolipidni monosloj i uronjen je u lipidnu jezgru lipoproteina. Istovremeno se oslobađa mjesto za holesterol u fosfolipidnom sloju, koji se može napuniti kolesterolom iz ćelijskih membrana ili iz drugih lipoproteina. Stoga se čini da je HDL zamka holesterola kao rezultat djelovanja LHAT-a.
Sinteza žučnih kiselina
U jetri se dio holesterola pretvara u žučne kiseline. Galne kiseline se mogu smatrati derivatima holanske kiseline (Sl. 10.32).
Kolanska kiselina kao takva ne nastaje u tijelu. U hepatocitima, holesterol direktno stvara čejnodeoksiholičnu i količnu kiselinu - primarne žučne kiseline (Sl. 10.33, vidi takođe Sl. 10.12).
Biosinteza holesterola
Biosinteza holesterola se javlja u endoplazmatskom retikuluu. Izvor svih atoma ugljika u molekuli je acetil-SCoA, koji dolazi iz mitohondrija kao dio citrata, kao u sintezi masnih kiselina. Biosinteza holesterola troši 18 ATP molekula i 13 NADPH molekula.
Stvaranje holesterola se događa u više od 30 reakcija, koje se mogu grupirati u više faza.
1. Sinteza mevalonske kiseline.
Prve dvije reakcije sinteze podudaraju se s reakcijama ketogeneze, ali nakon sinteze 3-hidroksi-3-metilglutarila-ScoA, enzim ulazi hidroksimetil-glutaril-ScoA reduktaza (HMG-SCOA reduktaza), formirajući mevalonsku kiselinu.
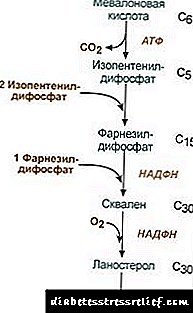 |
Šema reakcionih shema sinteze holesterola2. Sinteza izopentenil difosfata. U ovoj fazi se na mevalonsku kiselinu vežu tri ostatka fosfata, zatim se dekarboksilira i dehidrira. 3. Nakon kombiniranja tri molekule izopentenil-difosfata, sintetizira se farnezil-difosfat. 4. Sinteza skvalena dolazi kada se vežu dva ostatka farnesil-difosfata. 5. Nakon složenih reakcija, linearni skvalen ciklizira u lanosterol. 6. Uklanjanje viška metilnih grupa, obnavljanje i izomerizacija molekula dovodi do pojave holesterola. Regulacija aktivnosti hidroksimetilglutaril-S-CoA reduktaze3. Brzina biosinteze holesterola takođe ovisi o koncentraciji specifični proteinski nosačosiguravajući vezivanje i transport hidrofobnih metabolizama intermedijarne sinteze. Možete pitati ili ostaviti svoje mišljenje. Glavna tačka regulacije je reakcija stvaranja mevalonske kiseline. 1. Alosterična regulacija. Holesterol, a u jetri - i žučne kiseline inhibiraju HMG-CoA reduktazu. 2. Represija sinteze holesterola HMG-CoA reduktaze. 3. Regulacija fosforilacijom-deposforilacijom HMG-CoA reduktaze, aktivni nefosforilirani oblik. Glukagon izaziva deaktivaciju, a inzulin izaziva aktivaciju kroz složenu kaskadu reakcija. Tako se brzina sinteze holesterola mijenja s promjenom stanja apsorpcije i postabsorpcije. 4. Brzina sinteze HMG-CoA reduktaze u jetri podložna je jasnim dnevnim fluktuacijama: maksimalna u ponoć, a minimalna ujutro. Izmjena holesterola U ćelijama Esterifikacija holesterola nastaje kada je izložena acil-CoA-holesterol-aciltransferaza (AHAT): Acyl-CoA + holesterol ® NS-CoA + Acylchoterol U ćelijama se formira uglavnom linoleilhoterol. Esteri se nalaze uglavnom u citosolu kao dio kapljica lipida. Stvaranje estera može se s jedne strane smatrati mehanizmom za uklanjanje viška holesterola iz membrana, a s druge strane, kao mehanizmom za skladištenje holesterola u ćeliji. Mobilizacija rezervi odvija se uz sudjelovanje enzima esterazahidroliziranje estera holesterola: Acylchoterol + H2O ® masna kiselina + holesterol Sinteza i hidroliza estera je posebno aktivna u stanicama nadbubrežne kore. U krvi lipoproteini formiranje estera događa se učešćem lecitin-holesterol-aciltransferaza (LHAT), katalizirajući prenos acilnog ostatka iz lecitina u holesterol. LHAT nastaje u jetri, izlučuje se u krvotok i veže se za lipoproteine. Najaktivniji je LHAT u HDL-u gdje je lokaliziran u površinskom sloju. Ovdje formirani holesteroli su hidrofobni i uronjeni su u lipidno jezgro. U fosfolipidnom monolastu ima slobodnog prostora za holesterol koji se može ispuniti holesterolom iz ćelijskih membrana ili iz drugih lipoproteina. Stoga se čini da je HDL zamka holesterola kao rezultat djelovanja LHAT-a. Sinteza žučnih kiselina U jetri se dio holesterola pretvara u žučne kiseline. Žučne kiseline se mogu smatrati derivatima holanske kiseline. Kolanska kiselina kao takva ne nastaje u tijelu. U hepatocitima iz holesterola nastaju primarne žučne kiseline - chenodeoxycholic i holik. Nakon izlučivanja žuči u crijeva pod djelovanjem enzima crevne flore, iz njih se stvaraju sekundarne žučne kiseline - litološke i deoxycholic. Apsorbiraju se iz creva, s tim da krv portalne vene ulazi u jetru, a potom u žuč. Žuća sadrži uglavnom konjugirane žučne kiseline, tj. Njihove spojeve s glicinom ili taurinom. Koncentracija žučnih kiselina u žuči iznosi oko 1%. Glavni dio žučnih kiselina je uključen u hepatoenteric cirkulacija.Mali dio žučnih kiselina - oko 0,5 g dnevno - izlučuje se iz izmeta. To se nadoknađuje sintezom novih žučnih kiselina u jetri, fond žučnih kiselina se ažurira u otprilike 10 dana. Holesterol se izlučuje uglavnom kroz crijeva. U creva ulazi sa hranom i iz jetre kao deo žuči. Holesterol koji se apsorbira u krv sadrži frakciju koja potiče iz žuči (endogeni holesterolsintetizira se u jetri), a frakcija dobivena iz hrane (egzogeni holesterol) Odstranjivanje holesterola iz tkiva nastaje njegovom oksidacijom u žučnim kiselinama u jetri, nakon čega slijedi njihovo izlučivanje izmetom (otprilike 0,5 g dnevno) i izlučivanjem nepromijenjenog kolesterola (također s izmetom). U stacionarnom stanju: (Holesterolkraj + Holesterolex) - (holesterolizlučuju + Žučne kiselineizlučuju) = 0 Ako se ova ravnoteža poremeti, mijenja se koncentracija holesterola u tkivima i u krvi. Povećani holesterol u krvi - hiperholesterolemija. To povećava verovatnoću ateroskleroze i žučnih kamenaca. LIPIDNI PROPIS REGULACIJE Metabolizam lipida reguliše centralni nervni sistem. Dugotrajan negativan emocionalni stres, povećanje ispuštanja kateholamina u krvotok može uzrokovati primjetni gubitak težine. Akcija glukagon na lipolitički sistem sličan je djelovanju kateholamina. Adrenalin i norepinefrin povećavaju aktivnost tkivne lipaze i brzinu lipolize u masnom tkivu, kao rezultat, povećava se sadržaj masnih kiselina u krvnoj plazmi. Inzulin ima suprotan učinak adrenalina i glukagona na lipolizu i mobilizaciju masnih kiselina. Hormon rasta stimulira lipolizu, inducirajući sintezu acenilat ciklaze. Hipofunkcija hipofize dovodi do taloženja masti u tijelu (pretilost hipofize). Tiroksin, spolni hormonitakođe utiču na metabolizam lipida. Uklanjanje spolnih žlijezda kod životinja uzrokuje taloženje viška masnoće. LIPIDNI METABOLSKI POREMEĆAJI Holesterol je steroid specifičan za životinjske organizme. Glavno mesto njegovog formiranja u ljudskom telu je jetra gde se 50% holesterola sintetiše, 15–20% formira u tankom crevu, ostatak se sintetiše u koži, nadbubrežnoj kore i žlezdi. Izvori formiranja holesterolskog fonda i načini njegovog trošenja prikazani su na slici 22.1. Sl. 22.1. Stvaranje i distribucija holesterola u tijelu. Holesterol u ljudskom tijelu (ukupna količina od oko 140 g) može se uvjetno podijeliti u tri bazena: 30 g) koji se brzo razmenjuje, sastoji se od holesterola crevne stijenke, krvne plazme, jetre i drugih parenhimskih organa, obnavljanje dolazi za 30 dana (1 g / dan), 50 g), polako izmjenjujući holesterol drugih organa i tkiva, 60 g), vrlo sporo izmjenjenog kolesterola kičmene moždine i mozga, vezivnog tkiva, stopa ažuriranja izračunava se tijekom godina. Sinteza holesterola se događa u citosolu ćelija. Ovo je jedan od najdužih metaboličkih puteva u ljudskom tijelu. Nastaje u 3 stupnja: prvi završava stvaranjem mevalonske kiseline, drugi tvorbom skvalena (linearna ugljikovodična struktura koja se sastoji od 30 atoma ugljika). Tokom treće faze skvalen se pretvara u molekulu lanosterola, tada dolazi do 20 uzastopnih reakcija koje pretvaraju lanosterol u holesterol. U nekim tkivima hidroksilna grupa holesterola esterificira i formira estere. Reakcija je katalizirana unutarćelijskim enzimom AHAT (acylCoA: holesterola aciltransferaza). Reakcija esterifikacije se u krvi događa i kod HDL-a, gdje se nalazi enzim LHAT (lecitin: holesterola aciltransferaza). Esteri holesterola su oblik u kojem se transportuju krvlju ili deponuju u ćelijama. U krvi se oko 75% holesterola nalazi u obliku estera. Sinteza kolesterola se regulira utjecajem na aktivnost i količinu ključnog enzima procesa - 3-hidroksi-3-metilglutaril-CoA reduktaze (HMG-CoA reduktaza). To se postiže na dva načina: 1. Fosforilacija / defosforilacija HMG-CoA reduktaze. Inzulin potiče defosforilaciju HMG-CoA reduktaze, prevodeći je u aktivno stanje. Zbog toga se u apsorpcijskom periodu povećava sinteza holesterola. Tokom ovog perioda, raspoloživost polaznog supstrata za sintezu, acetil-CoA, takođe se povećava. Glukagon ima suprotan učinak: putem proteinske kinaze A potiče fosforilaciju HMG-CoA reduktaze, pretvarajući je u neaktivno stanje. Kao rezultat toga, inhibira se sinteza holesterola u periodu postabsorpcije i tokom posta. 2. Inhibicija sinteze HMG-CoA reduktaze.Holesterol (krajnji produkt metaboličkog puta) smanjuje brzinu transkripcije gena HMG-CoA reduktaze, čime inhibira vlastitu sintezu, a žučne kiseline također izazivaju sličan učinak. Transport holesterola u krvi vrši se kao dio lijeka. LP-ovi obezbeđuju egzogeni holesterol u tkivima, određuju njegov protok između organa i izlučivanje iz organizma. Egzogeni holesterol se dostavlja u jetru kao dio zaostalog ChM. Tamo zajedno sa sintetizovanim endogenim holesterolom tvori zajednički fond. U hepatocitima se TAG i holesterol pakuju u VLDL i u tom obliku se izlučuju u krv. U krvi se VLDL pod djelovanjem LP-lipaze, koja hidrolizira TAG u glicerol i masne kiseline, prvo pretvara u LSPP, a potom u LDL koji sadrži do 55% kolesterola i njegovih estera. LDL je glavni transportni oblik holesterola u kojem se dostavlja tkivima (70% kolesterola i njegovih estera u krvi je dio LDL). LDL iz krvi ulazi u jetru (do 75%) i druga tkiva koja na svojoj površini imaju LDL receptore. Ako količina kolesterola koji uđe u ćeliju premašuje njegove potrebe, tada se smanjuje sinteza LDL receptora, što smanjuje protok holesterola iz krvi. S padom koncentracije slobodnog kolesterola u ćeliji, naprotiv, aktivira se sinteza receptora. Hormoni su uključeni u regulaciju sinteze LDL receptora: inzulin, trijodtironin i spolni hormoni povećavaju stvaranje receptora, a glukokortikoidi smanjuju. U takozvanom "prijevozu kolesterola u povratku", tj. put koji osigurava povratak holesterola u jetru, HDL igra glavnu ulogu. Sintetišu se u jetri u obliku nezrelih prekursora koji praktično ne sadrže holesterol i TAG. HDL prekursori u krvi zasićeni su holesterolom, primajući ih iz drugih LP-a i staničnih membrana. Prijenos kolesterola u HDL uključuje enzim LHAT smješten na njihovoj površini. Ovaj enzim veže ostatak masne kiseline iz fosfatidilholina (lecitina) i holesterola. Kao rezultat toga, formira se hidrofobna molekula estera holesterola koja se kreće unutar HDL-a. Tako se ne pijani HDL, obogaćeni holesterolom, pretvaraju u HDL 3 - zrele i veće čestice. HDL 3 izmjenjuju estere kolesterola za TAG sadržane u VLDL i STD uz sudjelovanje specifičnog proteina koji prenosi estere holesterola između lipoproteina. U ovom slučaju HDL 3 pretvaraju u HDL2, čija se veličina povećava zbog nakupljanja TAG-a. VLDL i STDL pod utjecajem LP-lipaze pretvaraju se u LDL, koji uglavnom dostavljaju holesterol jetri. Mali dio kolesterola dostavlja se u jetru HDL2 i HDL. Sinteza žučnih kiselina. U jetri se iz holesterola dnevno sintetiše 500–700 mg žučnih kiselina. Njihovo stvaranje uključuje reakcije uvođenja hidroksilnih skupina uz sudjelovanje hidroksilaza i djelomičnu oksidaciju bočnog lanca holesterola (Sl. 22.2): Sl. 22.2. Šema stvaranja žučnih kiselina. Prva reakcija sinteze - stvaranje 7-a-hidroksihoterola - regulatorna je. Aktivnost enzima koji katalizira ovu reakciju inhibira krajnji produkt puta, žučne kiseline. Drugi mehanizam regulacije je fosforilacija / defosforilacija enzima (fosforilirani oblik 7-a-hidroksilaze je aktivan). Regulacija je moguća i promjenom količine enzima: holesterol potiče transkripciju gena 7-a-hidroksilaze, a žučne kiseline potiskuju. Hormoni štitnjače induciraju sintezu 7-a-hidroksilaze i represiju estrogena. Takav utjecaj estrogena na sintezu žučnih kiselina objašnjava zašto se žučna bolest javlja kod žena 3-4 puta češće nego kod muškaraca. Količne i kenoodeoksične kiseline nastale iz holesterola nazivaju se "primarnim žučnim kiselinama". Većina tih kiselina podvrgava se konjugaciji - dodavanju molekula glicina ili taurina u karboksilnu grupu žučne kiseline. Konjugacija započinje stvaranjem aktivnog oblika žučnih kiselina - derivata CoA, zatim se vežu taurin ili glicin, i kao rezultat nastaju 4 varijante konjugata: tauroholična i taurohenodeoksiholična, glikoholična i glikohenodeoksiholična kiselina. Oni su značajno jači emulgatori od originalnih žučnih kiselina. Konjugati s glicinom formiraju se 3 puta više nego sa taurinom, pošto je količina taurina u tijelu ograničena. U crijevima se mala količina konjugata primarnih žučnih kiselina pod djelovanjem bakterijskih enzima pretvara u sekundarne žučne kiseline. Deoksiholična kiselina, koja se formira iz holika, i litoholik, koja nastaje iz deoksiholične kiseline, manje su rastvorljiva i sporije se apsorbuju u crevima. Oko 95% žučnih kiselina koje uđu u crijevo vraća se u jetru putem portalne vene, a zatim se ponovo izlučuju u žuč i ponovno se koriste u emulgiranju masti. Taj put žučnih kiselina naziva se enterohepatička cirkulacija. Sa izmetom se uglavnom uklanjaju sekundarne žučne kiseline. Žučna kamena bolest (kolelitijaza) je patološki proces u kojem se formiraju kamenje u žučnoj kesici, čija je osnova holesterol. Oslobađanje holesterola iz žuči mora biti praćeno proporcionalnim oslobađanjem žučnih kiselina i fosfolipida koji zadržavaju hidrofobne molekule holesterola u micelarnom stanju. Razlozi koji vode do promjene omjera žučnih kiselina i kolesterola u žuči jesu: hrana bogata holesterolom, visokokalorična prehrana, stagnacija žuči u žučnom mjehuru, poremećena enterohepatička cirkulacija, poremećena sinteza žučnih kiselina, infekcija žučnog mjehura. Kod većine bolesnika sa kolelitijazom se povećava sinteza kolesterola, a sinteza žučnih kiselina iz njega se usporava, što dovodi do nesrazmjernog broja kolesterola i žučnih kiselina koje se izlučuju u žuč. Kao rezultat toga, holesterol se počinje taložiti u žučnoj kesici, tvoreći viskozni talog koji se postupno stvrdnjava. Ponekad je impregniran bilirubinom, proteinima i kalcijevim solima. Kamenje se može sastojati samo od holesterola (holesterola od kamenja) ili mješavine holesterola, bilirubina, proteina i kalcijuma. Kamen za holesterol obično je bijel, a mješoviti kamenje smeđe boje u različitim nijansama. U početnoj fazi stvaranja kamena, kenoodeoksikiselinska kiselina može se koristiti kao lijek. Jednom u žučnoj kesici postepeno rastvara kamenje holesterola, ali ovo je spor proces koji traje nekoliko mjeseci. Ateroskleroza je patologija koju karakterizira pojava aterogenih plakova na unutrašnjoj površini vaskularnog zida. Jedan od glavnih razloga razvoja takve patologije jeste kršenje ravnoteže između unosa holesterola iz hrane, njegove sinteze i izlučivanja iz organizma. Pacijenti s aterosklerozom imaju povišene koncentracije LDL i VLDL. Postoji inverzna veza između koncentracije HDL i vjerovatnoće za razvoj ateroskleroze. Ovo je u skladu sa konceptom funkcionisanja LDL kao prenositelja holesterola u tkivima, i HDL iz tkiva. Osnovni metabolički "preduvjet" za razvoj ateroskleroze je hiperholesterolemija. (visok holesterol u krvi). Hiperholesterolemija se razvija: 1. zbog prekomjernog unosa holesterola, ugljikohidrata i masti, 2. genetska predispozicija koja se sastoji od nasljednih oštećenja u strukturi receptora LDL ili apoB-100, kao i u povećanoj sintezi ili izlučivanju apoB-100 (u slučaju porodične kombinirane hiperlipidemije, u kojoj su povišene koncentracije kolesterola i kolesterola u krvi). Važnu ulogu u mehanizmima razvoja ateroskleroze igra modifikacija lijeka. Promjene u normalnoj strukturi lipida i proteina u LDL čine ih stranim u tijelu i zato su dostupnije za hvatanje fagocitima. Modifikacija lijeka može se dogoditi kroz nekoliko mehanizama: 1. glikozilacija proteina do koje dolazi kada se koncentracija glukoze u krvi poveća, 2. modifikacija peroksida, što dovodi do promjena u lipidima lipoproteina i strukture apoB-100, 3. stvaranje autoimunih kompleksa antitela LP (izmenjeni lekovi mogu izazvati stvaranje autoantitijela). Modifikovani LDL apsorbuju se makrofagi. Ovaj proces nije regulisan količinom apsorbovane holesterola, kao u slučaju njegovog ulaska u ćelije preko specifičnih receptora, zbog čega su makrofagi preopterećeni holesterolom i pretvaraju se u „pjenaste ćelije“ koje prodiru u subendotelni prostor. To dovodi do stvaranja lipidnih mrlja ili pruga u zidu krvnih žila. U ovoj fazi vaskularni endotel može održavati svoju strukturu. Sa porastom broja penastih ćelija dolazi do oštećenja endotela. Oštećenja doprinose aktiviranju trombocita. Kao rezultat, oni luče tromboksan, koji stimulira agregaciju trombocita, a također počinju stvarati faktor rasta koji potiče iz trombocita, a koji stimulira proliferaciju stanica glatkih mišića. Potonji migriraju iz medijalnog u unutarnji sloj arterijskog zida i tako doprinose rastu plaka. Nadalje, plak raste s vlaknastim tkivom, stanice ispod vlaknaste membrane su nekrotične, a holesterol se taloži u međućelijskom prostoru. U posljednjim fazama razvoja, plak je impregniran kalcijevim solima i postaje vrlo gust. U području plaka često se formiraju ugrušci krvi koji blokiraju lumen žile što dovodi do akutnog zatajenja krvotoka na odgovarajućem mjestu tkiva i do nastanka srčanog udara. Regulacija sinteze holesterola - njegovog ključnog enzima (HMG-CoA reduktaza), provodi se na različite načine. Fosforilacija / defosforilacija HMG reduktaze. S porastom omjera inzulin / glukagon, ovaj enzim fosforilira i prelazi u aktivno stanje. Delovanje inzulina se izvodi preko 2 enzima. HMG-CoA reduktaza kinaza fosfataza, koja kinazu pretvara u neaktivno defosforilirano stanje: Fosfotaza HMG-CoA reduktaza pretvarajući je u defosforilirano aktivno stanje. Rezultat tih reakcija je stvaranje defosforiliranog aktivnog oblika HMG-CoA reduktaze. Zbog toga se tokom perioda apsorpcije holesterol povećava. Tokom tog razdoblja, povećava se i dostupnost početnog supstrata za sintezu holesterola - acetil - CoA (kao rezultat konzumiranja hrane koja sadrži ugljikohidrate i masti. Kako se tijekom razgradnje glukoze i masnih kiselina formira acetil CoA). U postabsorbentnom stanju glukagon putem proteingenaze A stimulira fosforilaciju HMG - CoA - reduktaze, pretvarajući je u neaktivno stanje. Ovo je djelovanje pojačano činjenicom da istodobno glukagon potiče fosforilaciju i inaktivaciju fosfotaze HMG-CoA reduktaze, zadržavajući na taj način HMG-CoA reduktazu u fosforiliranom neaktivnom stanju. Kao rezultat toga, endogena sinteza inhibira sintezu holesterola u periodu postabsorpcije i tokom posta. Ako je udio kolesterola u hrani doveden na 2%, tada se sinteza endogenog kolesterola naglo smanjila. Ali potpuni prestanak sinteze holesterola ne dolazi. Stepen inhibicije biosinteze holesterola pod uticajem holesterola koji dolazi iz hrane varira od osobe do osobe. Ovo ukazuje na individualnost procesa stvaranja holesterola. Smanjenjem intenziteta sinteze holesterola moguće je smanjiti njegovu koncentraciju u krvi. Ako se poremeti ravnoteža između unosa holesterola s hranom i njegove sinteze u tijelu s jedne strane, i izlučivanja žučnih kiselina i holesterola s druge strane, mijenja se koncentracija kolesterola u tkivima i krvi. Najozbiljnije posljedice povezane su s povećanjem koncentracije kolesterola u krvi (hiperholesterolemija), dok vjerovatnoća za razvoj ateroskleroze i holelitiaze raste. Porodična hiperholesterolemija (HCS) - ovaj je oblik mnogo češći - oko 1 pacijenta na 200 ljudi. Nasljedna manjkavost HCS-a je kršenje apsorpcije LDL-a ćelijama, a samim tim i smanjenje stope katabolizma LDL-a. Kao rezultat toga, koncentracija LDL u krvi raste, kao i holesterol jer ga ima puno u LDL. Stoga je za HCS karakteristično taloženje holesterola u tkivima, posebno u koži (ksantomima), u zidovima arterija. Inhibicija sinteze HMG-CoA reduktaze Konačni produkt metaboličkog puta holesterola. Smanjuje brzinu transkripcije gena HMG-CoA reduktaze, čime inhibira vlastitu sintezu. Jetra aktivno sintetizira žučne kiseline iz kolesterola, te stoga žučne kiseline inhibiraju aktivnost gena HMG-CoA reduktaze. Budući da HMG-CoA reduktaza postoji nakon sinteze oko 3, inhibicija sinteze enzima ovog holesterola je efikasna regulacija. Pogledajte video: Regulacija Aktivnosti Gena - Lac Operon (Maj 2024).
|